The use of alternative fertilizers offers a promising approach to improving soil health and crop productivity, yet their relative effects on rhizosphere microbial communities remain insufficiently understood. This two-year field study on the Huang-Huai Plain, China, assessed the impacts of controlled-release fertilizer (CRF), organic fertilizer (OF), and microbial fertilizer (MF), relative to a conventional synthetic fertilizer (CF), on soil properties, wheat yield, and microbial community composition. Soil chemical parameters, microbial diversity (via 16S rRNA and ITS sequencing), and wheat yield were analyzed. OF significantly enhanced soil organic matter (14.97%), available nitrogen (28.70%), phosphorus (20.59%), potassium (33.06%), and grain yield (17.58%) compared to CF, likely due to sustained nutrient release and stimulation of microbial activity. In contrast, CRF decreased soil organic matter (−19.2%) and phosphorus availability, with only modest yield improvement (3.50%). MF enriched plant-beneficial taxa, including Bacillus and Arthrobacter, and improved yield by 9.39%. Fungal communities showed greater responsiveness to fertilizer type than bacterial communities, with OF and CRF promoting notable increases in fungal diversity. LEfSe analysis revealed treatment-specific microbial biomarkers such as Saccharothrix (OF), Azotobacter (CRF), and Nitrospira (MF), while correlation analysis linked Cyphellophora (OF) and Epicoccum (CRF) to yield enhancement. These findings underscore the potential of organic amendments to simultaneously boost soil fertility, microbial diversity, and crop productivity, outperforming controlled-release and microbial fertilizers. MF demonstrated promise for microbiome-targeted interventions, whereas CRF may pose risks to long-term soil health. This study supports microbiome-informed, organic-inclusive fertilization strategies for sustainable agriculture.
| Published in | International Journal of Applied Agricultural Sciences (Volume 11, Issue 4) |
| DOI | 10.11648/j.ijaas.20251104.11 |
| Page(s) | 101-119 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
New Fertilizers, Rhizosphere Microbiome, Winter Wheat, Yield, Soil Fertility
CRF | Controlled-Release Fertilizer |
OF | Organic Fertilizer |
MF | Microbial Fertilizer |
SOM | Soil Organic Matter |
TN | Total Nitrogen |
AN | Available Nitrogen |
AP | Available Phosphorus |
AK | Available Potassium |
RCBD | Randomized Complete Block Design |
Kg | Kilogram |
Ha | Hectare |
CFU | Colony Forming Unit |
Cm | Centimeter |
w/v | Weight to Volume |
PCR | Polymerase Chain Reaction |
OTU | Operational Taxonomic Unit |
μL | Microliter |
PCoA | Principal Coordinates Analysis |
LEfSe | LDA Effect Size |
LDA | Linear Discriminant Analysis |
ANOVA | ||||||
|---|---|---|---|---|---|---|
Sum of squares | freedom | mean square | F | significance | ||
SOM | Inter group | 176.133 | 3.000 | 58.711 | 46.993 | 0.000 |
within-subjects | 9.995 | 8.000 | 1.249 | |||
total | 186.127 | 11.000 | ||||
pH | Inter group | 0.175 | 3.000 | 0.058 | 8.404 | 0.007 |
within-subjects | 0.056 | 8.000 | 0.007 | |||
total | 0.231 | 11.000 | ||||
AN | Inter group | 2699.083 | 3.000 | 899.694 | 11.910 | 0.003 |
within-subjects | 604.333 | 8.000 | 75.542 | |||
total | 3303.417 | 11.000 | ||||
AP | Inter group | 8933.801 | 3.000 | 2977.934 | 10.414 | 0.004 |
within-subjects | 2287.724 | 8.000 | 285.966 | |||
total | 11221.526 | 11.000 | ||||
AK | Inter group | 19267.254 | 3.000 | 6422.418 | 26.733 | 0.000 |
within-subjects | 1921.925 | 8.000 | 240.241 | |||
total | 21189.179 | 11.000 | ||||
TN | Inter group | 0.026 | 3.000 | 0.009 | 0.682 | 0.587 |
within-subjects | 0.103 | 8.000 | 0.013 | |||
total | 0.129 | 11.000 | ||||
Yield | Inter group | 3.015 | 3.000 | 1.005 | 13.376 | 0.002 |
within-subjects | 0.601 | 8.000 | 0.075 | |||
total | 3.616 | 11.000 | ||||
| [1] | Bohra, A., Choudhary, M., Bennett, D., Joshi, R., Mir, R. R., & Varshney, R. K. (2024). Drought-tolerant wheat for enhancing global food security. Functional & integrative genomics, 24(6), 212. |
| [2] | Erenstein, O., Jaleta, M., Mottaleb, K. A., Sonder, K., Donovan, J., Braun, HJ. (2022). Global Trends in Wheat Production, Consumption and Trade. In: Reynolds, M. P., Braun, HJ. (eds) Wheat Improvement. Springer, Cham. |
| [3] | GRIGORAS Mircea Adrian, POPESCU Agatha, PAMFIL Doru, HAS Ioan & GIDEA Mihai. (2012). Conservation Agriculture versus Conventional Agriculture: The Influence of Agriculture System, Fertilization and Plant Protection on Wheat Yield. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 40(1), 188-188. |
| [4] | Priya E., Sudipta Sarkar, Pradip K. Maji, A review on slow-release fertilizer: Nutrient release mechanism and agricultural sustainability, Journal of Environmental Chemical Engineering, Volume 12, Issue 4, 2024, 113211, ISSN 2213-3437, |
| [5] | Tadvani, H. K., Bijanzadeh, E., & Najafi-Ghiri, M. (2024). Synergistic effect of salicylic acid and biochar on biochemical properties, yield and nutrient uptake of triticale under water stress. Heliyon, 10(17), e37152. |
| [6] | Donghao Xu, Gerard H. Ros, Qichao Zhu, Fusuo Zhang & Wim de Vries. (2025). Spatial optimization of manure and fertilizer application strategies to minimize nutrient surpluses and acidification rates in croplands of a typical Chinese county. Journal of Cleaner Production, 503, 145401-145401. |
| [7] | Bhardwaj, A. K., Rajwar, D., Yadav, R. K., Chaudhari, S. K., & Sharma, D. K. (2021). Nitrogen Availability and Use Efficiency in Wheat Crop as Influenced by the Organic-Input Quality Under Major Integrated Nutrient Management Systems. Frontiers in plant science, 12, 634448. |
| [8] | Wang, R., Yang, L., Guo, M., Lin, X., Wang, R., & Guo, S. (2024). Effects of microplastic properties and dissolved organic matter on phosphorus availability in soil and aqueous mediums. Environmental pollution (Barking, Essex: 1987), 340(Pt 2), 122784. |
| [9] | Liu, X., Rong, X., Yang, J., Li, H., Hu, W., Yang, Y., Jiang, G., Xiao, R., Deng, X., Xie, G., Luo, G., & Zhang, J. (2023). Community succession of microbial populations related to CNPS biological transformations regulates product maturity during cow-manure-driven composting. Bioresource technology, 369, 128493. |
| [10] | Xiang, Y., Liu, Y., Gong, M., Tong, Y., Liu, Y., Zhao, G., & Yang, J. (2023). Preparation of Novel Biodegradable Polymer Slow-Release Fertilizers to Improve Nutrient Release Performance and Soil Phosphorus Availability. Polymers, 15(10), 2242. |
| [11] | Jindo, K., Audette, Y., Olivares, F. L., Canellas, L. P., Smith, D. S., & Paul Voroney, R. (2023). Biotic and abiotic effects of soil organic matter on the phytoavailable phosphorus in soils: a review. Chemical and biological technologies in agriculture, 10(1), 29. |
| [12] | Li, K., Rahman, S. U., Rehman, A., Li, H., Hui, N., & Khalid, M. Shaping rhizocompartments and phyllosphere microbiomes and antibiotic resistance genes: The influence of different fertilizer regimes and biochar application. Journal of Hazardous Materials. 2025, 487, 137148. |
| [13] | Li, P., Tian, Y., Yang, K., Tian, M., Zhu, Y., Chen, X., Hu, R., Qin, T., Liu, Y., Peng, S., Yi, Z., Liu, Z., Ao, H., & Li, J. (2024). Mechanism of microbial action of the inoculated nitrogen-fixing bacterium for growth promotion and yield enhancement in rice (Oryza sativa L.). Advanced biotechnology, 2(4), 32. |
| [14] | Zhang, J., Liu, Q., Wang, D., & Zhang, Z. (2023). Soil Microbial Community, Soil Quality, and Productivity along a Chronosequence of Larix principis-rupprechtii Forests. Plants (Basel, Switzerland), 12(16), 2913. |
| [15] | Frac, M., Panek, J., Gryta, A., Oszust, K., Pertile, G., Siegieda, D.,... & Pietramellara, G. Legume-cereal intercropping as a strategy of regenerative agriculture supporting reverse of biodiversity loss-relevance of microbiome-based research. In ARPHA Conference Abstracts (No. 2). Pensoft Publishers. 2023. |
| [16] | Lagunas, B., Richards, L., Sergaki, C., Burgess, J., Pardal, A. J., Hussain, R. M. F., Richmond, B. L., Baxter, L., Roy, P., Pakidi, A., Stovold, G., Vázquez, S., Ott, S., Schäfer, P., & Gifford, M. L. (2023). Rhizobial nitrogen fixation efficiency shapes endosphere bacterial communities and Medicago truncatula host growth. Microbiome, 11(1), 146. |
| [17] | Duan, Y., Chen, R., Zhang, R., Jiang, W., Chen, X., Yin, C., & Mao, Z. (2021). Isolation, Identification, and Antibacterial Mechanisms of Bacillus amyloliquefaciens QSB-6 and Its Effect on Plant Roots. Frontiers in microbiology, 12, 746799. |
| [18] | Perkowski, E. A., Terrones, J., German, H. L., & Smith, N. G. (2024). Symbiotic nitrogen fixation reduces belowground biomass carbon costs of nitrogen acquisition under low, but not high, nitrogen availability. AoB PLANTS, 16(5), plae051. |
| [19] | Lin, W., Lin, M., Zhou, H., Wu, H., Li, Z., & Lin, W. The effects of chemical and organic fertilizer usage on rhizosphere soil in tea orchards. PLoS ONE. 2019, 14(5), e0217018. |
| [20] | Bourak, K., Sare, A. R., Allaoui, A., Jijakli, M. H., & Massart, S. Impact of two phosphorus fertilizer formulations on wheat physiology, rhizosphere, and rhizoplane microbiota. International Journal of Molecular Sciences. 2023, 24(12), 9879. |
| [21] | Nelson, Darrell W., and Lee E. Sommers. "Total carbon, organic carbon, and organic matter." Methods of soil analysis: Part 3 Chemical methods 5, 1996. 961-1010. |
| [22] | Bremner, John M. "Nitrogen‐total." Methods of soil analysis: Part 3 Chemical methods 5, 1996. 1085-1121. |
| [23] | Olsen, Sterling Robertson. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. No. 939. US Department of Agriculture, 1954. |
| [24] | Roberts, T. L.; Ross, W. J.; Norman, R. J.; Slaton, N. A.; Wilson, C. E. Predicting nitrogen fertilizer needs for rice in Arkansas using alkaline Hydrolyzable-Nitrogen. Soil Science Society of America Journal, 2011. 75 (3), 1161-1171. |
| [25] | Wang, T.; Xu, J.; Chen, J.; Liu, P.; Hou, X.; Yang, L.; Zhang, L. Progress in microbial fertilizer regulation of crop growth and soil remediation research. Plants 2024, 13 (3), 346. |
| [26] | Shu, X.; He, J.; Zhou, Z.; Xia, L.; Hu, Y.; Zhang, Y.; Zhang, Y.; Luo, Y.; Chu, H.; Liu, W.; Yuan, S.; Gao, X.; Wang, C. Organic amendments enhance soil microbial diversity, microbial functionality and crop yields: A meta-analysis. The Science of the Total Environment 2022, 829, 154627. |
| [27] | Ondrasek, G.; Begić, H. B.; Zovko, M.; Filipović, L.; Meriño-Gergichevich, C.; Savić, R.; Rengel, Z. Biogeochemistry of soil organic matter in agro ecosystems & environmental implications. The Science of the Total Environment 2018, 658, 1559-1573. |
| [28] | Zhang, S., Ren, T., Fang, Y. et al. Enhancing soil labile organic matter through oilseed rape-rice rotation and straw returning in paddy-upland systems. Plant Soil (2025). |
| [29] | Li, R.; Gao, Y.; Chen, Q.; Li, Z.; Gao, F.; Meng, Q.; Li, T.; Liu, A.; Wang, Q.; Wu, L.; Wang, Y.; Liu, Z.; Zhang, M. Blended controlled-release nitrogen fertilizer with straw returning improved soil nitrogen availability, soil microbial community, and root morphology of wheat. Soil and Tillage Research 2021, 212, 105045. |
| [30] | Chae, H. G.; Margenot, A. J.; Jeon, J.-R.; Kim, M. S.; Jang, K.-S.; Yoon, H. Y.; Kim, P. J.; Lee, J. G. Linking the humification of organic amendments with size aggregate distribution: Insights into molecular composition using FT-ICR-MS. The Science of the Total Environment 2024, 927, 172147. |
| [31] | Wang, C.; Kuzyakov, Y. Soil organic matter priming: The pH effects. Global Change Biology 2024, 30 (6). |
| [32] | Heuchan, S. M.; Fan, B.; Kowalski, J. J.; Gillies, E. R.; Henry, H. a. L. Development of Fertilizer Coatings from Polyglyoxylate-Polyester Blends Responsive to Root-Driven pH Change. Journal of Agricultural and Food Chemistry 2019, 67 (46), 12720-12729. |
| [33] | Baddar, Z. E.; Unrine, J. M. Effects of Soil pH and Coatings on the Efficacy of Polymer coated ZnO Nanoparticulate fertilizers in Wheat (Triticum aestivum). Environmental Science & Technology 2021, 55 (20), 13532-13540. |
| [34] | Xu, J.; Mohamed, E.; Li, Q.; Lu, T.; Yu, H.; Jiang, W. Effect of humic acid addition on buffering capacity and nutrient storage capacity of soilless substrates. Frontiers in Plant Science 2021, 12. |
| [35] | Wambacq, E.; Alloul, A.; Grunert, O.; Carrette, J.; Vermeir, P.; Spanoghe, J.; Sakarika, M.; Vlaeminck, S. E.; Haesaert, G. Aerobes and phototrophs as microbial organic fertilizers: Exploring mineralization, fertilization and plant protection features. PLoS ONE 2022, 17 (2), e0262497. |
| [36] | Lori, M.; Symnaczik, S.; Mäder, P.; De Deyn, G.; Gattinger, A. Organic farming enhances soil microbial abundance and activity—A meta-analysis and meta-regression. PLoS ONE 2017, 12 (7), e0180442. |
| [37] | Xing, X.; Xu, H.; Wang, D.; Yang, X.; Qin, H.; Zhu, B. Nitrogen use aggravates bacterial diversity and network complexity responses to temperature. Scientific Reports 2022, 12 (1). |
| [38] | Shade, A. Microbiome rescue: directing resilience of environmental microbial communities. Current Opinion in Microbiology 2023, 72, 102263. |
| [39] | Sabir, M. S.; Shahzadi, F.; Ali, F.; Shakeela, Q.; Niaz, Z.; Ahmed, S. Comparative Effect of Fertilization Practices on Soil Microbial Diversity and Activity: An Overview. Current Microbiology 2021, 78 (10), 3644-3655. |
| [40] | Niu, B.; Fu, G. Response of plant diversity and soil microbial diversity to warming and increased precipitation in alpine grasslands on the Qinghai-Xizang Plateau - A review. The Science of the Total Environment 2023, 912, 168878. |
| [41] | Robinson, A.; Babinski, M.; Xu, Y.; Kelliher, J.; Longley, R.; Chain, P. A centralized resource for bacterial-fungal interactions research. Fungal Biology 2023, 127 (5), 1005-1009. |
| [42] | Khanna, K.; Kohli, S. K.; Sharma, N.; Kour, J.; Devi, K.; Bhardwaj, T.; Dhiman, S.; Singh, A. D.; Sharma, N.; Sharma, A.; Ohri, P.; Bhardwaj, R.; Ahmad, P.; Alam, P.; Albalawi, T. H. Phytomicrobiome communications: Novel implications for stress resistance in plants. Frontiers in Microbiology 2022, 13. |
| [43] | Kalwani, M.; Chakdar, H.; Srivastava, A.; Pabbi, S.; Shukla, P. Effects of nanofertilizers on soil and plant-associated microbial communities: Emerging trends and perspectives. Chemosphere 2021, 287, 132107. |
| [44] | Li, L.; Huang, D.; Hu, Y.; Rudling, N. M.; Canniffe, D. P.; Wang, F.; Wang, Y. Globally distributed Myxococcota with photosynthesis gene clusters illuminate the origin and evolution of a potentially chimeric lifestyle. Nature Communications 2023, 14 (1). |
| [45] | Pardue, E. J.; Sartorio, M. G.; Jana, B.; Scott, N. E.; Beatty, W. L.; Ortiz-Marquez, J. C.; Van Opijnen, T.; Hsu, F.-F.; Potter, R. F.; Feldman, M. F. Dual membrane-spanning anti-sigma factors regulate vesiculation in Bacteroides thetaiotaomicron. Proceedings of the National Academy of Sciences 2024, 121 (10). |
| [46] | Mujakić, I.; Piwosz, K.; Koblížek, M. Phylum gemmatimonadota and its role in the environment. Microorganisms 2022, 10 (1), 151. |
| [47] | Luo, L.; Zhao, C.; Wang, E.; Raza, A.; Yin, C. Bacillus amyloliquefaciens as an excellent agent for biofertilizer and biocontrol in agriculture: An overview for its mechanisms. Microbiological Research 2022, 259, 127016. |
| [48] | Kong, X.; Bai, Z.; Jin, T.; Jin, D.; Pan, J.; Yu, X.; Cernava, T. Arthrobacter is a universal responder to di-n-butyl phthalate (DBP) contamination in soils from various geographical locations. Journal of Hazardous Materials 2021, 422, 126914. |
| [49] | Kuzikova, I. L.; Medvedeva, N. G. Long-Chain alkylphenol biodegradation potential of soil ascomycota. Doklady Biological Sciences 2023, 511 (1), 228-234. |
| [50] | Yang, Z.; Dong, H.; Zhang, S.; Jiang, J.; Zhu, H.; Yang, H.; Li, L. Isolation and identification of mycorrhizal helper bacteria of Vaccinium uliginosum and their interaction with mycorrhizal fungi. Frontiers in Microbiology 2023, 14. |
| [51] | Sonnabend, R.; Seiler, L.; Gressler, M. Regulation of the Leucine Metabolism in Mortierella alpina. Journal of Fungi 2022, 8 (2), 196. |
| [52] | Wei, B.; Luo, X.; Zhou, Z.-Y.; Hu, G.-A.; Li, L.; Lin, H.-W.; Wang, H. Discovering the secondary metabolic potential of Saccharothrix. Biotechnology Advances 2023, 70, 108295. |
| [53] | Das, H. K. Azotobacters as biofertilizer. Advances in Applied Microbiology 2019, 1-43. |
| [54] | Wu, Y.; Li, H.; Li, Y. Biomineralization Induced by Cells of Sporosarcina pasteurii: Mechanisms, Applications and Challenges. Microorganisms 2021, 9 (11), 2396. |
| [55] | Al-Ajeel, S.; Spasov, E.; Sauder, L. A.; McKnight, M. M.; Neufeld, J. D. Ammonia-oxidizing archaea and complete ammonia-oxidizing Nitrospira in water treatment systems. Water Research X 2022, 15, 100131. |
| [56] | Pellegrinetti, T. A.; De Cássia Mesquita Da Cunha, I.; De Chaves, M. G.; De Freitas, A. S.; Passos, G. S.; Da Silva, A. V. R.; Cotta, S. R.; Tsai, S. M.; Mendes, L. W. Genomic insights of Fictibacillus terranigra sp. nov., a versatile metabolic bacterium from Amazonian Dark Earths. Brazilian Journal of Microbiology 2024, 55 (2), 1817-1828. |
| [57] | Liu, W.; Fan, Y.; He, P.; Chen, H. Complete genome sequence of a nitrate reducing bacteria, Algoriphagus sp. Y33 isolated from the water of the Indian Ocean. Marine Genomics 2021, 59, 100861. |
| [58] | Torres-Garcia, D.; Gené, J.; García, D.; Cano-Lira, J. F. Insights into Some Onygenalean Fungi from Freshwater Sediments in Spain and Description of Novel Taxa. Journal of Fungi 2023, 9 (12), 1129. |
| [59] | Chang, L.; Lu, H.; Chen, H.; Tang, X.; Zhao, J.; Zhang, H.; Chen, Y. Q.; Chen, W. Lipid metabolism research in oleaginous fungus Mortierella alpina: Current progress and future prospects. Biotechnology Advances 2021, 54, 107794. |
| [60] | Abed, R. M. Exploring fungal biodiversity of genus epicoccum and their biotechnological potential. In Fungal biology; 2021; pp 237-276. |
| [61] | Zhang, J.; Luo, D.; Li, C.; Wang, T.; Zhu, Z.; Huang, X. Advances and Prospects of Modern Processes of Coprophilous Fungal Spores: Applications in Paleoecological Studies. Advances in Earth Science, 2024. 39(5), 489-503. |
| [62] | Santana, J. D. S.; Custódio, F. A.; De Oliveira, J. A.; Pereira, O. L. Cyphellophora denticulata sp. nov. a new dark septate endophyte from banana roots in Brazil. Brazilian Journal of Microbiology 2025. |
| [63] | Luo, C.; He, Y.; Chen, Y. Rhizosphere Microbiome regulation: unlocking the potential for plant growth. Current Research in Microbial Sciences 2024, 8, 100322. |
APA Style
Wen, R., Cao, Y., Mohamed, S. M. R., Qihang, Y., Bo, W., et al. (2025). Different New Fertilizers Differentially Modulate Wheat Yield, Rhizosphere Microbiota and Soil Fertility. International Journal of Applied Agricultural Sciences, 11(4), 101-119. https://doi.org/10.11648/j.ijaas.20251104.11
ACS Style
Wen, R.; Cao, Y.; Mohamed, S. M. R.; Qihang, Y.; Bo, W., et al. Different New Fertilizers Differentially Modulate Wheat Yield, Rhizosphere Microbiota and Soil Fertility. Int. J. Appl. Agric. Sci. 2025, 11(4), 101-119. doi: 10.11648/j.ijaas.20251104.11
@article{10.11648/j.ijaas.20251104.11,
author = {Ran Wen and Yang Cao and Sohaila Mohamed Ramadan Mohamed and Yu Qihang and Wang Bo and Liu Hecheng},
title = {Different New Fertilizers Differentially Modulate Wheat Yield, Rhizosphere Microbiota and Soil Fertility
},
journal = {International Journal of Applied Agricultural Sciences},
volume = {11},
number = {4},
pages = {101-119},
doi = {10.11648/j.ijaas.20251104.11},
url = {https://doi.org/10.11648/j.ijaas.20251104.11},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ijaas.20251104.11},
abstract = {The use of alternative fertilizers offers a promising approach to improving soil health and crop productivity, yet their relative effects on rhizosphere microbial communities remain insufficiently understood. This two-year field study on the Huang-Huai Plain, China, assessed the impacts of controlled-release fertilizer (CRF), organic fertilizer (OF), and microbial fertilizer (MF), relative to a conventional synthetic fertilizer (CF), on soil properties, wheat yield, and microbial community composition. Soil chemical parameters, microbial diversity (via 16S rRNA and ITS sequencing), and wheat yield were analyzed. OF significantly enhanced soil organic matter (14.97%), available nitrogen (28.70%), phosphorus (20.59%), potassium (33.06%), and grain yield (17.58%) compared to CF, likely due to sustained nutrient release and stimulation of microbial activity. In contrast, CRF decreased soil organic matter (−19.2%) and phosphorus availability, with only modest yield improvement (3.50%). MF enriched plant-beneficial taxa, including Bacillus and Arthrobacter, and improved yield by 9.39%. Fungal communities showed greater responsiveness to fertilizer type than bacterial communities, with OF and CRF promoting notable increases in fungal diversity. LEfSe analysis revealed treatment-specific microbial biomarkers such as Saccharothrix (OF), Azotobacter (CRF), and Nitrospira (MF), while correlation analysis linked Cyphellophora (OF) and Epicoccum (CRF) to yield enhancement. These findings underscore the potential of organic amendments to simultaneously boost soil fertility, microbial diversity, and crop productivity, outperforming controlled-release and microbial fertilizers. MF demonstrated promise for microbiome-targeted interventions, whereas CRF may pose risks to long-term soil health. This study supports microbiome-informed, organic-inclusive fertilization strategies for sustainable agriculture.},
year = {2025}
}
TY - JOUR T1 - Different New Fertilizers Differentially Modulate Wheat Yield, Rhizosphere Microbiota and Soil Fertility AU - Ran Wen AU - Yang Cao AU - Sohaila Mohamed Ramadan Mohamed AU - Yu Qihang AU - Wang Bo AU - Liu Hecheng Y1 - 2025/07/14 PY - 2025 N1 - https://doi.org/10.11648/j.ijaas.20251104.11 DO - 10.11648/j.ijaas.20251104.11 T2 - International Journal of Applied Agricultural Sciences JF - International Journal of Applied Agricultural Sciences JO - International Journal of Applied Agricultural Sciences SP - 101 EP - 119 PB - Science Publishing Group SN - 2469-7885 UR - https://doi.org/10.11648/j.ijaas.20251104.11 AB - The use of alternative fertilizers offers a promising approach to improving soil health and crop productivity, yet their relative effects on rhizosphere microbial communities remain insufficiently understood. This two-year field study on the Huang-Huai Plain, China, assessed the impacts of controlled-release fertilizer (CRF), organic fertilizer (OF), and microbial fertilizer (MF), relative to a conventional synthetic fertilizer (CF), on soil properties, wheat yield, and microbial community composition. Soil chemical parameters, microbial diversity (via 16S rRNA and ITS sequencing), and wheat yield were analyzed. OF significantly enhanced soil organic matter (14.97%), available nitrogen (28.70%), phosphorus (20.59%), potassium (33.06%), and grain yield (17.58%) compared to CF, likely due to sustained nutrient release and stimulation of microbial activity. In contrast, CRF decreased soil organic matter (−19.2%) and phosphorus availability, with only modest yield improvement (3.50%). MF enriched plant-beneficial taxa, including Bacillus and Arthrobacter, and improved yield by 9.39%. Fungal communities showed greater responsiveness to fertilizer type than bacterial communities, with OF and CRF promoting notable increases in fungal diversity. LEfSe analysis revealed treatment-specific microbial biomarkers such as Saccharothrix (OF), Azotobacter (CRF), and Nitrospira (MF), while correlation analysis linked Cyphellophora (OF) and Epicoccum (CRF) to yield enhancement. These findings underscore the potential of organic amendments to simultaneously boost soil fertility, microbial diversity, and crop productivity, outperforming controlled-release and microbial fertilizers. MF demonstrated promise for microbiome-targeted interventions, whereas CRF may pose risks to long-term soil health. This study supports microbiome-informed, organic-inclusive fertilization strategies for sustainable agriculture. VL - 11 IS - 4 ER -
State Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping/College of Agronomy, Henan Agricultural University, Zhengzhou, China
Biography: Ran Wen is an undergraduate student enrolled in 2022 at the College of Agronomy, Henan Agricultural University, majoring in Agronomy (Shaoqui Innovation Experimental Class of New Agricultural Sciences), has achieved notable accomplishments as a core participant: a national silver award and a bronze award at the China International College Students Innovation Competition, a national bronze award at the Chuang Qing chun competition, and a first prize in the Central China Division of the National Agricultural Science Students Competition. Additionally, as the project leader, she presided over the 2023 Henan Agricultural University College Students Entrepreneurship Practice Program. Currently, mainly explores the fields of corn cultivation physiology and the regulation of crop stress resistance.
Research Fields: Microorganism, Ecological environment, Maize cultivation, maize physiological stress and regulation, soil properties

State Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping/College of Agronomy, Henan Agricultural University, Zhengzhou, China
Biography: Yang Cao is an undergraduate student (Class of 2022) majoring in Seed Science and Engineering at the College of Agronomy, Henan Agricultural University, specializing in Crop Cultivation and Farming Systems. Currently engaged in research practice at the Henan Pro-vincial Key Laboratory of Crop Physiology and Ecology, he is systematically studying the theories and technologies of high-yield and high-efficiency maize cultivation.
Research Fields: Microorganism, Ecological environment, Maize cultivation, maize physiological stress and regulation, soil properties

Department of Soil and Water Sciences, Faculty of Agriculture, Fayoum University, Fayoum, Egypt
Biography: Sohaila Mohamed Ramadan Mohamed, B.Sc. Student | Department of Soil and Water Sciences, Faculty of Agriculture, Fayoum Uni-versity, Egypt. Education: Expected B.Sc. in Land and Water Sciences (2025). Research Project: "Water Quality Assessment in Fayoum Governorate" Technical Skills: Field water sampling and laboratory analysis (spectrophotometry, titration), Water sampling techniques, Using laboratory equipment (pH meters, spectrophotometers), Data collection and report preparation Research Interests: Water resources, Soil management, Environmental sustainability.
Research Fields: Water resources-, Soil management,, Environmental sustainability

State Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping/College of Agronomy, Henan Agricultural University, Zhengzhou, China
Biography: Qihang Yu is an undergraduate student majoring in Agronomy at the College of Agronomy, Henan Agricultural University, enrolled in 2021. Currently, he focuses on maize stress cultivation research, assisting in the collection and statistical analysis of field phenotypic data. He is committed to integrating cutting-edge technologies with traditional agronomy to explore efficient and green development models for modern agriculture.
Research Fields: High temperature stress, drought stress, high temperature drought stress, soil properties, Stress Resistance Modulator Regulator Development

State Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping/College of Agronomy, Henan Agricultural University, Zhengzhou, China
Biography: Wang Bo is a 2023-level undergraduate student majoring in Agronomy at the College of Agronomy of Henan Agricultural University. She has taken core courses such as Molecular Biology and Genetics as her main subjects. She has accumulated solid foundation in field data modeling and application of agricultural information technology. Currently, she is conducting research on the physiological mechanism of high-yield corn cultivation and is responsible for data collection and precise fertilization in field experiments.
Research Fields: High temperature stress, drought stress, high temperature drought stress, soil properties, Stress Resistance Modulator Regulator Development

State Key Laboratory of High-Efficiency Production of Wheat-Maize Double Cropping/College of Agronomy, Henan Agricultural University, Zhengzhou, China
Biography: Hecheng Liu, an undergraduate student majoring in Agronomy at the College of Agriculture of Henan Agricultural University since 2021, was awarded the title of "Innovation Star" of the Third Henan Province University Students in 2024. He is an outstanding representative of the university who has consecutively won this honor for three years. Focusing on agricultural science and technology innovation, he integrates classroom learning with the demands of rural areas through participating in scientific research, technology development, and the promotion of achievements, dedicated to serving the development of agriculture, rural areas, and farmers. Currently, he mainly explores the fields of corn cultivation physiology and the regulation of crop stress resistance.
Research Fields: High temperature stress, drought stress, high temperature drought stress, soil properties, Stress Resistance Modulator Regulator Development

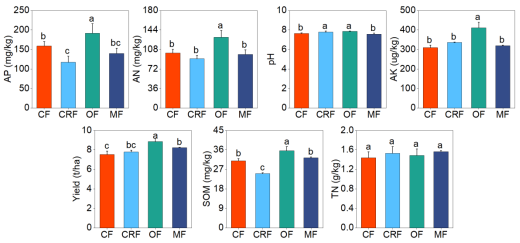
Figure 1. Effects of different fertilizers on soil chemical properties and wheat grain yield. A; Soil organic matter (SOM), B; Soil pH, C; Available nitrogen (AN), D; Available phosphorus (AP), E; Available potassium (AK), F; Total Nitrogen (TN), and G; Wheat grain yield. Results are shown for conventional (CF), controlled-release (CRF), organic (OF), and microbial (MF) fertilizers. Data are presented as mean ± standard error (n = 4 treatments * 3 replicates = 12). Columns with different lowercase letters are significantly different (p < 0.05).
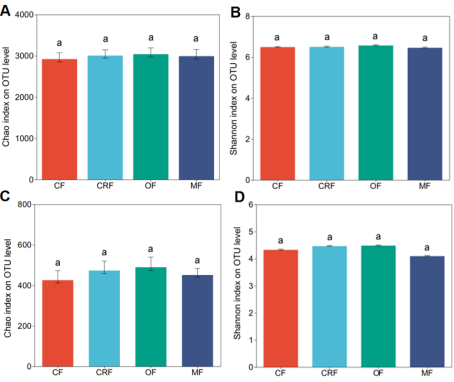
Figure 2. Effects of fertilizers on bacterial and fungal alpha diversity. Chao1 estimates microbial richness of (A) bacteria and (C) fungi. Shannon diversity index accounts for abundance and evenness in (B) bacteria and (D) fungi. Treatments: conventional (CF), controlled-release (CRF), organic (OF), and microbial (MF) fertilizers.
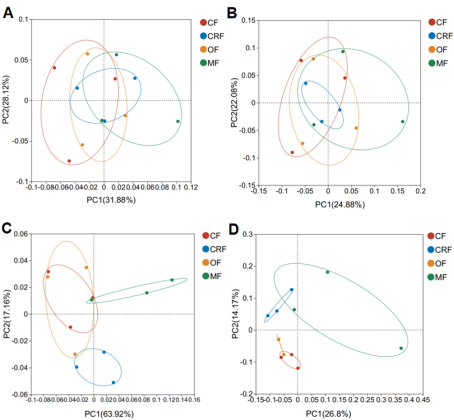
Figure 3. Principal Coordinate Analysis (PCoA) of bacterial and fungal beta diversity based on Bray-Curtis distances. Results are shown at phyla (A: bacteria, C: fungi) and genera (B: bacteria, D: fungi) levels. Ellipses represent 95% confidence intervals for each treatment group: conventional (CF), controlled-release (CRF), organic (OF), and microbial (MF) fertilizers.
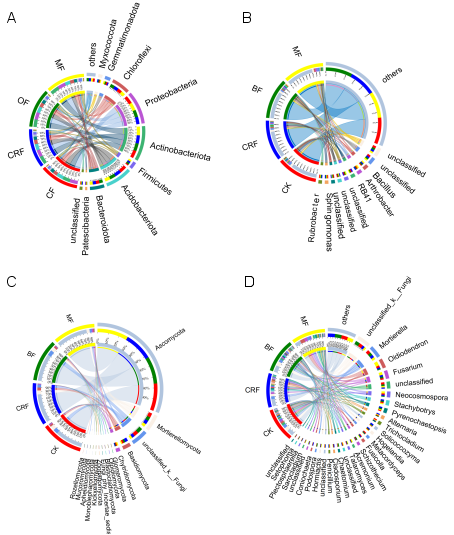
Figure 4. Circos diagrams showing the relative abundance of microbial taxa across fertilizer treatments. Results are shown at phyla (A: bacteria, C: fungi) and genera (B: bacteria, D: fungi) levels. In each diagram, the left semicircle represents treatment groups—conventional (CF), controlled-release (CRF), organic (OF), and microbial (MF) fertilizers— while the right semicircle represents microbial taxa, with outer ribbons for taxonomic groups and inner ribbons for treatments. Ribbon widths correspond to proportional abundances within each treatment.
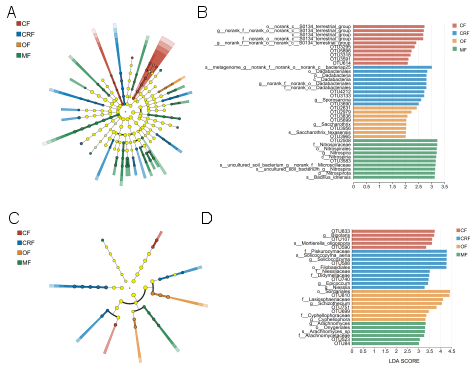
Figure 5. Composition of the bacterial and fungal community in the soil at the genus level. A; bacterial, C; fungal cladograms: Nodes represent taxa significantly enriched in treatments (colored) or non-significant taxa (light yellow). LEfSe identifies taxa driving differences. B; bacterial (LDA > 3), D; fungal (LDA > 2) bar charts. Treatments: conventional (CF), controlled-release (CRF), organic (OF), and microbial (MF) fertilizers.
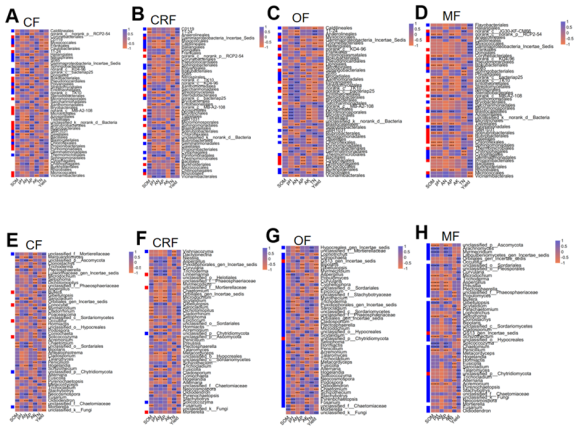
Figure 6. Heatmap of correlations between microbial genera and soil properties or wheat yield under different fertilizer treatments. (A-D) bacterial genera correlations. (E-H) fungal genera correlations. Rows (Y-axis) represent microbial genera; columns (X-axis) represent soil properties and yield. Correlation coefficients (r-values) are color-coded, with significance levels indicated (*p < 0.05, **p < 0.01, ***p < 0.001).
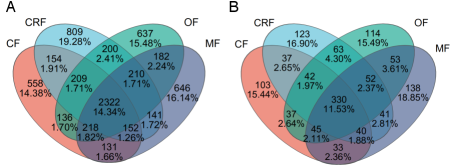
Figure A1. Venn diagram represents unique bacteria and fungi species under different alternative fertilizers (CF, CRF, OF, MF).
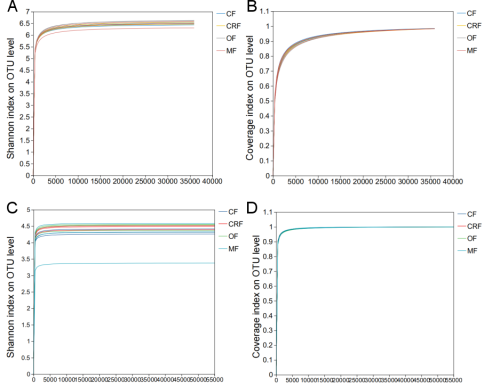
Figure A2. Dilution curve of community coverage and diversity index of soil bacteria and fungi at OTU level.
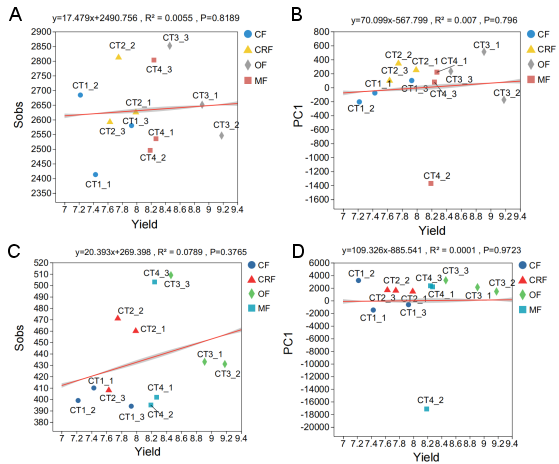
Figure A3. This figure represents the correlation between yield and microbiota structure, assessing the size of the impact on differences in sample community composition.

